La distylie de Primula vulgaris
favorise un transfert précis du pollen
entre morphes floraux

L'hétérostylie favorise la pollinisation intermorphique (assurée par des insectes) à cause de la position réciproque du stigmate et des anthères dans les fleurs brévistylées et longistylées. De plus, chez la plupart des espèces qui présentent une hétérostylie, un système d'auto-incompatibilité mettant en jeu des intéractions moléculaires limite fortement l'autofécondation et la fécondation intramorphique. . Les deux systèmes concourent à promouvoir la fécondation intermorphique.
Quelle est la part qui revient à l'hétérostylie?.
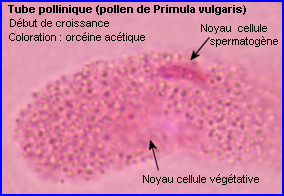
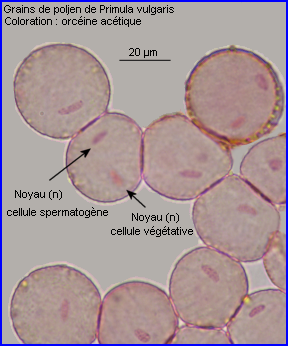
Les grains de pollen sont formés dans les anthères des étamines à partir des cellules mères qui subissent la méiose. A maturité, chaque grain de pollen de Primula vulgaris contient 2 cellules haploïdes (voir image), l'une d'entre-elles étant la cellule spermatogène. La couche la plus externe de la paroi du grain, l'exine, présente une ornementation avec des parties en creux dans lesquelles se retrouvent des protéines et glycoprotéines produites par le tapis de l'anthère lors de la maturation du pollen. Ces molécules correspondent à l'expression d'un génotype diploïde et certaines peuvent jouer le rôle de marqueurs du grain de pollen.
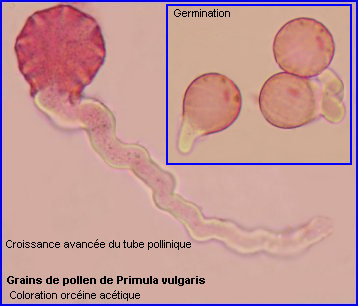
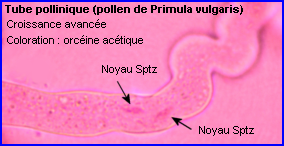
Pour que la double fécondation puisse avoir lieu, il faut qu'un grain de pollen livre 2 gamètes mâles au gamétophyte femelle (sac embryonnaire) situé dans un ovule donc dans l'ovaire. Il doit d'abord être réceptionné par un stigmate puis germer et développer un tube pollinique qui s'allonge dans le style, atteint l'ovaire puis un ovule et libère les 2 spermatozoïdes issus de la division de la cellule spermatogène. Il y a autopollinisation puis autofécondation quand le grain de pollen qui fournit les gamètes mâles et le pistil réceptif appartiennent à la même fleur ou à 2 fleurs du même pied sinon pollinisation et fécondation sont croisées.
Un système biochimique d'incompatibilité qui limite l'auto-pollinisation et la pollinisation intramorphique
Chez Primula vulgaris, l'autopollinisation et la pollinisation intramorphe sont le plus souvent infructueuses, soit par inhibition de la germination du pollen soit par interruption de la croissance du tube pollinique. Il y a donc incompatibilité entre le pollen produit par les anthères d'une fleur d'un morphe floral et un stigmate d'une fleur du même morphe. A l'origine de l'incompatibilité, il y a la reconnaissance par le stigmate réceptif (peut-être par le style) de molécules associées à l'exine du pollen. Ces marqueurs sont secrétés par le tapis, tissu diploïde de l'anthère. Les gènes impliqués dans le système d'incompatibilité sont liés à ceux qui gouvernent le dimorphisme floral avec lesquels ils constituent le supergène S, transmis comme un facteur mandélien. Le phénotype d'incompatibilité est défini par le génotype diploïde de la plante mère (incompatibilité sporophytique).
Il y aura donc le plus souvent fécondation
"thrum" x "pin".
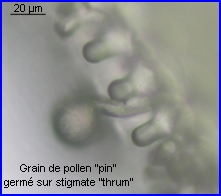
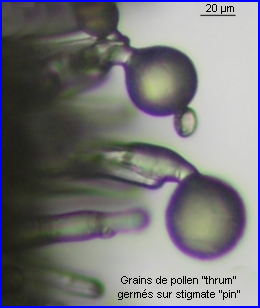
Le pollen des fleurs brévistylées et longistylées est réparti dans des zones différentes le long du proboscis et sur les poils de la tête d'un insecte qui récolte le nectar de Primula vulgaris. Le positionnement réciproque des anthères et des stigmates dans les fleurs des deux morphes floraux favorise la pollinisation intermorphique car le pollen d'un morphe se retrouve sur une partie de l'insecte qui correspond exactement à la zone qui touche le stigmate de l'autre morphe (voir ci-dessous).
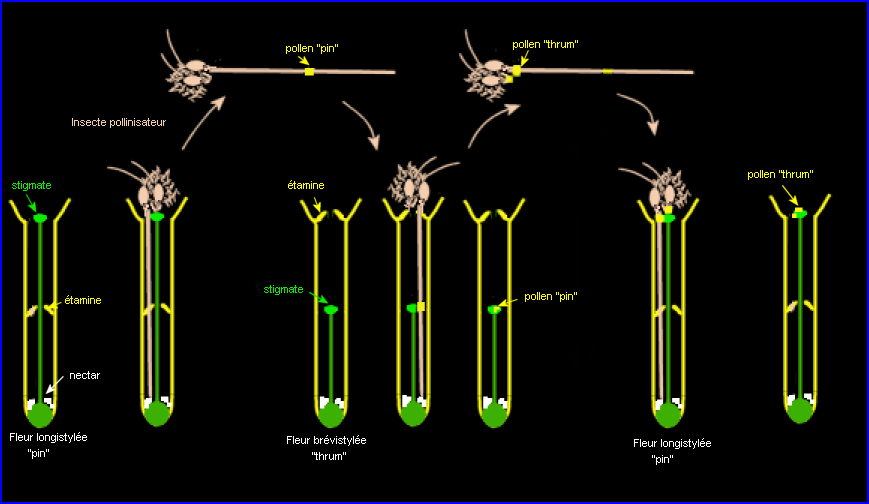
L'hétérostylie qui permet un transport intermorphe précis du pollen limiterait également l'interférence sexuelle au sein d'une même fleur.
Voir "Evolution des traits floraux et du mode de pollinisation des plantes à fleurs dans les zones de mégabiodiversité" de A. Quilichini.