
Le grain de pollen quiescent s'hydrate, in situ, au contact des papilles stigmatiques imprégnées d'exsudats gluants ou in vitro au contact d'un milieu convenablement préparé. Il devient alors métaboliquement très actif et une cyclose rotationnelle s'installe dans la celule végétative. Sous l'effet de la poussée de tugescence, il y a rupture de l'exine, formation puis allongement du tube pollinique, immédiatement parcouru par les courants cytoplasmiques qui matérialisent l'organisation fonctionnelle et stucturale polarisée du grain de pollen germé.
Objectifs : observer la cyclose dans les grains prêts à germer et dans les tubes polliniques, envisager son rôle dans la croissance du tube pollinique et dans le transport de l'unité germinale mâle.
Une étude des mécanismes moléculaires de la cyclose est proposée par ailleurs : voir "Cyclose dans le tube pollinique des Angiospermes" avec notamment un modèle de l'intervention du système actine-myosine-ATP.
Pour observer la cyclose, des grains de pollen bien hydratés sont montés dans un milieu qui les maintient en vie (vidéo). Ils sont ensuite colorés par l'orcéine acétique pour préciser leur organisation.
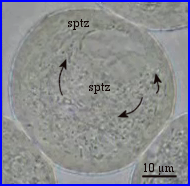
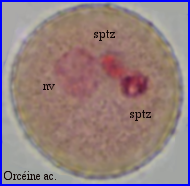
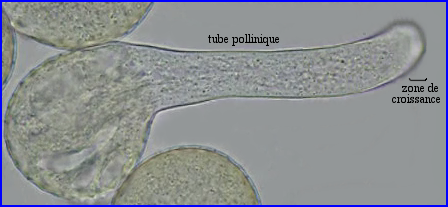
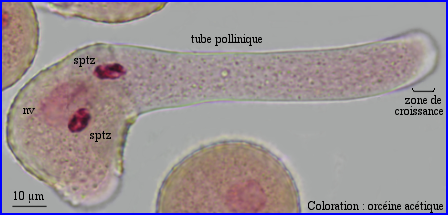
Le cytoplasme resté dans le grain s'est vacuolarisé.
La coloration à l'orcéine montre que l'unité germinale mâle n'est pas encore passée dans le tube pollinique.

Dans le grain, le vacuome occupe un volume important et les courants cytoplasmiques n'intéressent plus que de fines travées cytoplasmiques.

On peut évaluer la vitesse de déplacement d'une particule en la suivant le plus longtemps possible. La durée du déplacement est déterminée à l'aide de la base de temps de la vidéo et la distance parcourue est mesurée avec l'échelle reportée sur les images.
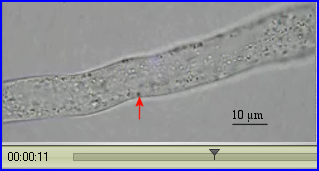
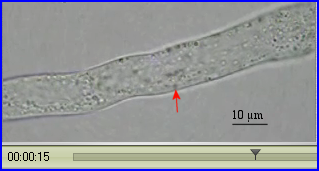
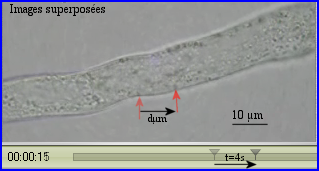
La reprise de l'activité métabolique d'un grain de pollen en contact avec des papilles stigmatiques récéptrices s'accompagne de la mise en mouvement de divers organites cytoplasmiques, RE, mitochondries, dictyosomes, vésicules golgiennes notamment. Dès que le grain de pollen germe, les courants cytoplasmiques sont repérables dans le tube pollinique. Ils favorisent les échanges de molécules à travers la membrane des organites et les rencontres entre enzymes et substrats. Ils sont indispensables pour permettre la croissance du tube qui a lieu dans la zone apicale claire. C'est le système moléculaire actine-myosine-ATP qui assure le trafic des organites. Ils sont pris en charge par la myosine qui se déplace le long des microfilaments (rails) d'actine polymérisée. La progression est rendue possible par la conversion d'une partie de l'énergie chimique de l'ATP en énergie de mouvement consécutivement à la formation des complexes d'actomyosine.
Ainsi, des vésicules de sécrétion d'origine golgienne contenant des précurseurs de la paroi (pectines, glycoprotéines diverses) sont amenées par le système actine-myosine-ATP au niveau de la zone apicale où elles échappent aux grands courants cytoplasmiques. Elles viennent alors fusionner avec la membrane plasmique déjà en place (création de nouvelle membrane) et leur contenu, libéré par exocytose, participe à l'édification de la nouvelle paroi.
En général, quand un tube atteint une certaine longueur, des dépôts successifs de callose (bouchons) isolent la partie en croissance qui demeure vivante des parties anciennes qui dégénèrent. La cyclose ne subsiste bien sûr que dans la zone vivante. Dans les tubes polliniques de l'Arum maculatum observés ici, il n'y a pas de bouchons de callose, il est vrai que leur longueur ne dépasse pas 350 µm ce qui représente environ 1/6 de la longueur moyenne totale qu'un tube doit atteindre pour parvenir, in situ, au niveau d'un ovule.
La vitesse de progression dans le tube de l'unité germinale mâle est plus lente que celle des courants cytoplasmiques. Les mécanismes moléculaires mis en jeu dans le transfert de l'ensemble noyau végétatif + 2 sptz sont probablement différents de ceux qui interviennent dans la cyclose. Ils ne sont pas élucidés. Des microtubules seraient impliqués.