Méiose dans le testicule de Locusta migratoria
Phénomènes chromosomiques
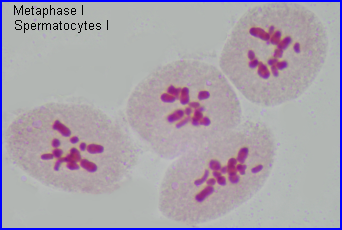
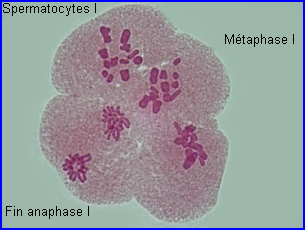
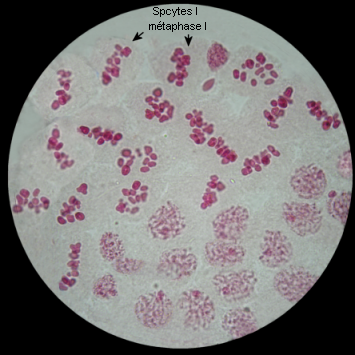
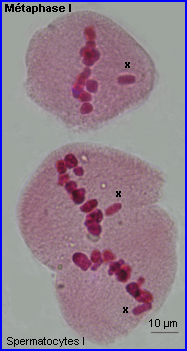
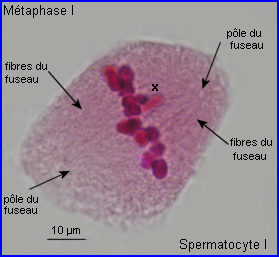
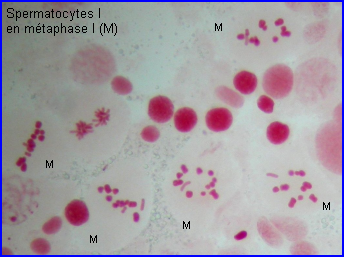
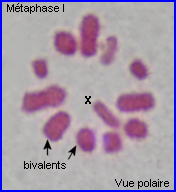
Métaphase I
A la métaphase, les bivalents (et X) sont placés dans le plan équatorial de la cellule par intervention des microtubules kinétochoriens.
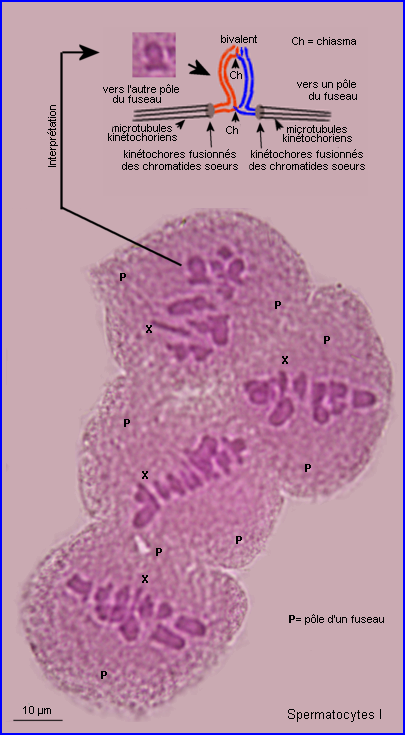
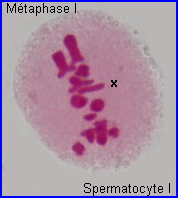
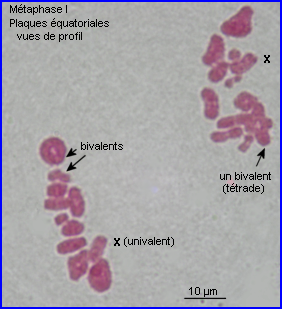
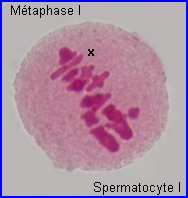
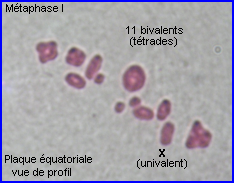
En plus de leur implication dans les recombinaisons génétiques (réassortiment des allèles des gènes) à la prophase I, les chiasmas ont un rôle déterminant dans la ségrégation des chromosomes homologues qui se prépare lors de la formation de la plaque équatoriale. En effet, leur maintien permet un positionnement interdépendant d'un chromosome et de son homologue sur le fuseau à la métaphase I, l'alignement des bivalents étant assuré par l'intervention des fibres fuseauriales.
Pour bien comprendre comment le positionnement des chomosomes à la métaphase I conditionne leur répartition à la phase suivante, il faut prendre en compte le mode d'intervention des microtubules kinétochoriens (non repérables) dans la mise en place des bivalents.
Les kinétochores (normalement un par chromatide) sont des assemblages protéiques associés à l'ADN centromérique auxquels sont attachés les microtubules (fibres) qui assurent la dynamique chromosomique lors des divisions cellulaires.
A la fin de la prophase I de méiose, les kinétochores des chromatides soeurs d'un chromosome apparaissent fusionnés en un seul complexe auquel sont liés des microtubules qui pointent tous vers le même pôle. Au sein d'un bivalent, les microtubules kinétochoriens de chaque chromosome homologue sont orientés en sens inverse. C'est par le jeu des tensions relatives exercées par les microtubules kinétochoriens de chacun des chromosomes homologues que le bivalent se retrouve positionné à la métaphase. En plaque équatoriale, l'orientation du kinétochore (donc du centromère) d'un des chromosomes du bivalent est opposée à celle de l'autre mais leur positionnement relatif est aléatoire (voir interprétation image centrale ci-dessus).